ФРУКТАНИ — полісахариди, побудовані із залишків D-фруктози; знайдені у вищих рослинах, зелених водоростях і бактеріях. Ф. або глюкофруктани накопичуються у рослинах у різних формах. Ф. — продукти трансфруктозилювання сахарози, тому кожна молекула містить один залишок D-глюкози, який не виявляє відновлювальних властивостей. В утворенні глікозидних зв’язків Ф. беруть участь тільки первинні групи –ОН. Молекула сахарози містить три первинні гідроксили, тому можливо утворення різних трисахаридів (рисунок), які є попередниками трьох типів Ф:
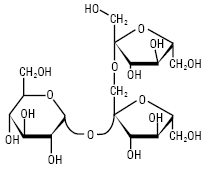
Ізокестоза (l-кестоза)
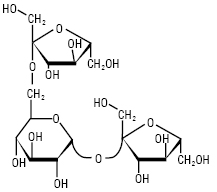
Кестоза
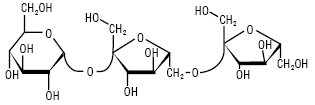
Неокестоза
Послідовне приєднання залишків D-фруктофуранози до 1-кестози зв’язками 2→1 дає інулін, а до фруктозного залишку 6-кестози зв’язками 2→6-флеїн (у рослинах) і леван (у бактеріях). Полісахариди на базі неокестози містять зв’язок 2→1 між залишками D-фруктофуранози, але зустрічаються рідко. Крім лінійних Ф., у рослинах поширені розгалужені форми Ф., в яких до головних ланцюгів інулінового типу приєднані бічні ланцюги зв’язками, характерними для флеїну, або до головних ланцюгів флеїнового типу приєднані бічні ланцюги зв’язками, характерними для інуліну. Біосинтез Ф. тісно пов’язаний з метаболізмом сахарози, яка служить як донором, так і акцептором залишків D-фруктофуранози. Ф. утворюють гомологічний ряд, кожний член якого відповідає основній формулі Г~ Ф~[Ф]n, де Г-Ф — цукрова група, [Ф] — залишок фруктофуранози. Гомологічний ряд Ф. починається з найнижчого гомолога сахарози (n=0), а не з фруктози. Наступним є трисахарид, далі йде тетрасахарид. Кожний попередній олігосахарид відрізняється від наступного на один залишок фруктози. Згідно з номенклатурою олігосахаридів вони називаються (фруктозил)-сахароза, (фруктозил)2-сахароза, (фруктозил)3-сахароза тощо. Вищим гомологом цього ряду є інулін, у якого n=35–40. Усі Ф. легкорозчиняються в гарячій воді. Для їх очищення використовують підвищену розчинність Ф. у водно-етанольних сумішах у порівнянні з іншими полісахаридами, осадження за допомогою Ва(ОН)2, а також хроматографічні методи. Ф. надзвичайно лабільні в кислому середовищі й руйнуються навіть при короткочасному нагріванні з розведеними (порядку 0,01 М) кислотами. Для встановлення хімічної будови Ф. використовують метод метилювання й ЯМР 13С. У клітинах рослин Ф. накопичуються у вакуолях і виконують роль резервного матеріалу (джерело фруктози), осморегулятора і антифризу. Вміст Ф. сягає 30% сухої маси в листках, а у спеціалізованих запасаючих органах (кореневищах, бульбах і цибулинах) може перевищувати 60%. Ф. — джерело енергії, яку рослини використовують у початковий період розвитку пагонів і листків або для відновлення нормального метаболізму після ушкодження рослини під час заморозків, поїдання тваринами або скошування. Порівняно низькомолекулярні Ф. (ступінь полімеризації не вище 100, зазвичай — 30–60) поширені у родині айстрових. Подібні Ф. продукують штами бактерій Streptococcus mutans. Більш високомолекулярні флеїни та Ф. змішаного типу (ступінь полімеризації до 300–400) характерні для однодольних рослин. Левани утворюються багатьма грампозитивними та грамнегативними бактеріями. Ф. не знайшли широкого практичного використання, але рослинна сировина, багата на Ф., може служити джерелом одержання D-фруктози. Рослинні Ф. типу інуліну виявляють імуномодулювальну, протипухлинну та протизапальну активність (див. Інулін).
Гудвин Т., Мерсер Э. Введение в биохимию растений. В 2 т. — М., 1986; Дудкин М.С. Введение в химию углеводов. — К., 1976; Кретович В.Л. Биохимия растений. — М., 1986; Химическая энциклопедия. В 5 т. / Гл. ред. И.Л. Кнунянц. — М., 1998.