
КОРІНЬ (лат. radix) — вегетативний, в основному підземний орган, характерний лише для вищих рослин; вперше сформувався у плаунів, хвощів, папоротей. К. багатьох видів рослин є цінною ЛРС, яка широко використовується у фармацевтичній промисловості для приготування різних видів ліків: настойок, таблеток, мазей тощо (валеріана лікарська, алтея лікарська, женьшень, ревінь, оман, дягель та ін.), а також у харчовій і цукровій промисловості, у тваринництві як кормові культури, у інших галузях промисловості: для отримання каучуку, гутаперчі, рослинних барвників. Усі ці продукти переробки широко використовуються у медицині та фармації. К. — це осьовий орган, який має циліндричну форму та радіальну симетрію. Основні функції К.: забезпечення мінерального та водного живлення (поглинання і транспорт води та мінеральних речовин); закріплення рослини у ґрунті; накопичення поживних речовин; синтез деяких речовин; вегетативне розмноження; симбіоз з бактеріями та зв’язок з іншими організмами ґрунту. Росте, в основному, кінчиком униз, виявляючи позитивний геотропізм. К. здатен рости доти, поки зберігається апікальна (верхівкова) меристема. Відрізняється від стебел тим, що не має листків, вузлів, міжвузлів, бруньок (за винятком придаткових), у нього відсутній інтеркалярний ріст, конус наростання захищений кореневим чохликом. Головний, або перший, К. насіннєвої рослини розвивається з зародкового корінця насінини. Від головного К. відходять бічні К. першого порядку, які закладаються у перициклі (шар клітин первинної меристеми). Вони дають початок бічним К. другого, третього та інших порядків. Сукупність К. однієї рослини — це коренева система. Кореневу систему, утворену головним та бічними К., називають стрижневою (рис. 1). Вона характерна для голонасінних та покритонасінних рослин класу дводольних. Існують придаткові К., які розвиваються від листків, стебел і старих К. Кореневу систему, утворену придатковими К., якщо головний К. припиняє ріст і зовні не відрізняється від інших або відмирає, називають мичкуватою. Вона характерна для рослин класу однодольних (жито, кукурудза, овес тощо). У деяких дводольних трав’янистих рослин коренева система мішана: досить добре розвинуті усі види К. (суниці, полуниці).

Рис.1. Кореневі системи і види К., їх видозміни: кореневі системи: 1 — стрижнева; 2 — мичкувата; видозміни коренів: 3 — коренеплід; 4 — коренебульби; 5 — К. з бульбочками; корені: 6 — водяні; 7 — повітряні; 8 — присоски, або гаусторії
К. і кореневі системи розвиваються в основному в ґрунті, але деякі рослини розвивають кореневу систему в повітрі (види орхідних), у воді (ряска, жабуриння), у товщі стебел інших рослин (присоски рослин-паразитів і напівпаразитів — омели, повитиці тощо). Повітряні К. за походженням придаткові й утворюють мичкувату кореневу систему. Можуть формуватися на пагонах рослин, рости вниз і, досягаючи ґрунту, вростати в нього та виконувати функцію звичайних К. Залежно від функцій К. можуть видозмінюватись. До метаморфозів К., що виконують запасальну функцію, належать коренеплоди (видозміна головного К.) та кореневі бульби або кореневі шишки (видозміна придаткових К.), у паренхімі яких відкладаються поживні речовини. Залежно від продуктів запасу і використання розрізняють 3 види коренеплодів: столові, які використовуються у харчуванні (столові буряки, морква, петрушка тощо); кормові, призначені для відгодівлі свійських тварин (кормові буряки, турнепс і т.п.); технічні, що використовуються у промисловості (цукрові буряки, кок-сагіз та ін.). Кореневі шишки (батат, чистяк) відрізняються від бульб — видозмінених пагонів (картоплі, топінамбуру) — тим, що не мають бічних бруньок (вічок) і лусок (видозмінених листків). Для мангрових рослин, які ростуть у зоні припливів, характерний розвиток ходульних К., які є додатковою опорою рослин. За походженням вони придаткові. Гаусторії — це придаткові всмоктувальні К. рослин-паразитів і напівпаразитів. Втягуючі, або контрактильні, К. здатні вкорочуватися та втягувати рослину або її частину (кореневище, цибулину) глибоко в ґрунт, що дозволяє легше переносити несприятливі умови (конюшина, проліски, женьшень, тюльпани тощо). Якщо на К. оселяється гриб, утворюється симбіотичний орган — грибокорінь (мікориза). Гіфи гриба заміняють рослині кореневі волоски. Для певних видів рослин характерна присутність певних видів грибів, які так і називають підосичник, підберезник, піддубник тощо. На К. рослин родин бобових, лохових утворюються бульбочки — розростання паренхімної тканини К. під дією ґрунтових бактерій, здатних фіксувати молекулярний азот атмосфери. Частина цього азоту утилізується рослиною. Цей симбіоз широко використовується у сільському господарстві для збагачення ґрунту азотистими речовинами.
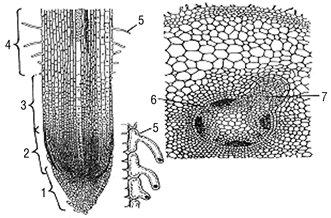
У кінчику К. на поздовжньому розрізі розрізняють чотири зони (рис. 2): ділення, росту, всмоктування, укріплення, які відрізняються за будовою та функціями.
а б
Рис. 2. Будова кінчика К. і утворення бічного К.: а — кінчик К.; б — утворення бічного К. із перициклу; 1 — кореневий чохлик; 2–4 — зони поділу клітин, росту і всмоктування; 5 — кореневий волосок; 6 — перицикл; 7 — бічний К.
До зони поділу клітин входить верхівка конуса наростання, де відбувається активне ділення клітин. Паренхімні клітини первинної (апікальної) меристеми цієї зони мають тонкі стінки, вкриті кореневим чохликом, який має трикутну форму, утворюються зовнішнім шаром апікальної меристеми, виділяють слиз, що сприяє просуванню К. у ґрунті. У водяних рослин кореневого чохлика немає. У зоні росту (2–5 мм) клітини меристеми збільшуються у розмірах, витягуються у довжину, поділ клітин поступово припиняється, що забезпечує ріст К. у довжину та просування його у ґрунті. У зоні всмоктування (5–20 мм) утворюються кореневі волоски — довгі вирости ризодерми, які збільшують всмоктувальну поверхню К., поглинають воду, розчини неорганічних речовин із ґрунту. Вони недовговічні, функціонують 10–20 діб, а потім відмирають. Кореневі волоски відсутні у водяних та болотяних рослин. У зоні всмоктування К. має первинну анатомічну будову. Зона укріплення в центральній частині характеризується утворенням бічних К. Укріплення рослин у ґрунті та пересування розчинів мінеральних речовин до надземних органів, а органічних — до клітин К. — це функція найдовшої зони К. У голонасінних та дводольних покритонасінних рослин у цій зоні К. має вторинну анатомічну будову, в однодольних — первинну, але відбуваються деякі зміни. Первинна будова К. (див. рис. 2–4) має три системи тканин: покривно-всмоктувальну з кореневими волосками епіблему, первинну кору та центральний циліндр. Епіблема виконує всмоктувальну та захисну функції. Первинна кора ділиться на екзодерму, мезодерму та ендодерму. Екзодерма складається з багатокутових, щільно зімкнутих клітин з частково зкорковілими оболонками, і виконує захисну та опорно-пропускну функції. Мезодерма — багатошарова запасальна частина, становить основну масу первинної кори, виконує транспортну та повітроносну функцію. Клітини живі, великі або багатокутові з тонкими або лігніфікованими оболонками (у однодольних), наповнені крохмальними зернами. Ендодерма — внутрішній однорядний шар первинної кори, що межує з центральним циліндром та виконує опорно-пропускну функцію. Центральний, або осьовий, циліндр займає серединну частину К., оточений кільцем перициклу, з якого формуються бічні К., а у голонасінних та дводольних — камбій та фелоген, які забезпечують вторинне потовщення К. Під перициклом розташований радіальний провідний пучок. У дводольних рослин променів ксилеми зазвичай не більше 6, а у однодольних — більше 6. У К. немає справжньої серцевини. У центрі можуть бути судини, склеренхіма, іноді — паренхіма, які утворюють несправжню серцевину. Вторинна будова К. формується в зоні укріплення голонасінних та дводольних покритонасінних рослин завдяки вторинним бічним меристемам — камбію та фелогену. Камбій утворює вторинну флоему та ксилему. Вторинна будова К. може бути пучкового (у деяких трав’янистих дводольних) (рис. 3) або непучкового типу (у дерев’янистих та деяких трав’янистих дводольних) (рис. 4). У дерев’янистих рослин К. зазвичай непучкового типу і у деревини формуються кільця річного приросту. Річне кільце — це сукупність тканин, утворених камбієм за один вегетаційний період. Особливістю коренеплодів є розростання запасної паренхіми у лубі (родина зонтичних) або у деревині (родина капустяних). У коренеплодах типу буряка (родина маревих) навколо центрального циліндра з безпучковою будовою внаслідок ділення клітин первинної флоеми та перициклу утворюються додаткові камбіальні кільця. У кореневих бульбах (жоржина, зозулинець, пшінка та ін.), як і у коренеплодів, добре розвинена запасна тканина.
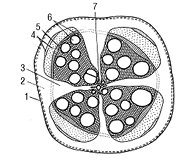
Рис. 3. Корінь дводольної трав’янистої рослини (пучковий тип): 1 — перидерма; 2 — корова паренхіма; 3 — серцевинні промені; відкритий колатеральний пучок: 4 — флоема первинна та вторинна; 5 — камбій; 6 — вторинна ксилема; 7 — первинна ксилема
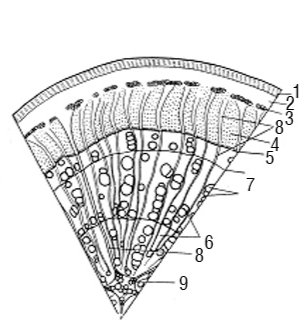
Рис. 4. Корінь безпучкового типу дерев’янистої рослини: 1 — перидерма; 2 — корова паренхіма; 3 — товстостінні елементи лубу — склереїди; 4 — тонкостінні елементи лубу; 5 — камбій; 6 — деревина; 7 — межа річного приросту; 8 — серцевинні промені; 9 — первинна ксилема
Діагностичною ознакою К., що відрізняє його від стебла, є наявність первинної ксилеми в центрі у вигляді зірочки з 2–5 променями.
Лотова Л.И. Морфология и анатомия высших растений. — М., 2001; Пронченко Г.Е. Лекарственные растительные средства / Под ред. А.П. Арзамасцева, И.А. Самилиной. — М., 2002; Сербін А.Г., Сіра Л.Н., Слободянюк Т.А. Фармацевтична ботаніка. — Вінниця, 2007; Эзау К. Анатомия семенных растений. В 2 т. — М.,1980.