
ЦИКЛІЧНІ НУКЛЕОТИДИ — сполуки, які утворюються при каталітичній дії циклаз нуклеозидтрифосфатів і перетворюють відповідні нуклеозидтрифосфати на їх циклічні форми шляхом відщеплення пірофосфату та замикання циклу між 3′- і 5′- вуглецевими атомами залишків α-D- рибофуранози. Серед Ц.н. найважливіші: циклічна-3′,5′- АМФ (3′,5′-цАМФ) та циклічна-3′,5′- гуанозинмонофосфорна кислота (3′,5′-цГМФ), які є регуляторами клітинного метаболізму:
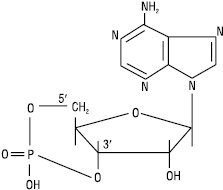
Циклічна 3′,5′-АМФ
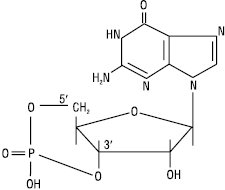
Циклічна 3′,5′-ГМФ
Шляхи біосинтезу цАМФ і цГМФ відрізняються та реалізуються через різні регуляторні системи, однак механізми їх впливу на клітинну активність подібні та зводяться до вибіркового фосфорилювання функціонально важливих клітинних білків. Обмін і функції Ц.н. у клітинах забезпечуються комплексом ферментів, які об’єднують в аденілат- і гуанілатциклазні системи, що містять специфічні ферменти синтезу: аденілат- і гуанілатциклази; ферменти перетворення Ц.н. — цАМФ- і цГМФ-фосфодіестерази; цАМФ- і цГМФ-залежні протеїнкінази та фосфопротеїнфосфатази, які усувають їх ефект. Аденілатциклаза каталізує утворення цАМФ з АТФ, а гуанілатциклаза каталізує утворення цГМФ з гуанозинтрифосфату (ГТФ).
Ц.н. відіграють в організмі роль вторинних посередників при передаванні гормонального імпульсу (сигналу). Їх регуляторна функція здійснюється внаслідок активації відповідних цГМФ- та цАМФ-залежних протеїнкіназ, які забезпечують каталітичну активність різних білків-ферментів. Фосфорильовані білки, що утворилися, викликають різні метаболічні ефекти. Функція одних білків після фосфорилювання їх протеїнкіназами активується, функція інших — пригнічується. Напр., під час дії адреналіну і глюкагону — гормонів, які підвищують рівень цАМФ у клітинах, активовані протеїнкінази фосфорилюють глікогенфосфорилазу, тобто переводять її з неактивної форми в активну та забезпечують розпад глікогену в печінці й скелетних м’язах (підвищують рівень глюкози в крові). У той же час внаслідок фосфорилювання відбувається інактивація ферменту глікогенсинтетази, що призводить до гальмування синтезу глікогену. Вплив інсуліну на внутрішньоклітинну утилізацію глюкози виявляється в стимуляції синтезу глікогену. Посилення глікогенезу в печінці та м’язах зумовлено зниженням рівня цАМФ внаслідок активації інсуліном фосфодіестерази. Зміна кількості цАМФ у клітині під впливом різних гормонів може здійснюватися як через аденілатциклазу, так і через фосфодіестеразу, яка каталізує гідроліз цАМФ з утворенням 5′-АМФ. Речовини, які гальмують фосфодіестеразу, проявляють ефект, подібний до дії гормону, але активація ферменту має протилежний ефект. Напр., інгібітори фосфодіестерази, похідні ксантинів: кофеїн, амінофілін, теофілін — підвищують рівень цАМФ, імітуючи ефект, який виявляють ендогенні гормони.
У клітинах-мішенях (напр. ендотеліальних клітинах) медіатор NO (оксид нітрогену) взаємодіє з іоном заліза активного центру гуанілатциклази, що призводить до швидкого утворення цГМФ. Підвищення концентрації цГМФ у клітинах гладких м’язів викликає активацію кіназ, що спричиняє розслаблення гладком’язових клітин судин та подальше їх розширення. Механізм дії оксиду нітрогену пояснює використання нітрогліцерину як ЛП для усунення гострого болю в серці, оскільки нітрогліцерин — джерело молекул NO, які й викликають розслаблення кровоносних судин та збільшення притоку крові в міокард. Ц.н. можуть виявляти синергічну або антагоністичну дію на регуляцію клітинного метаболізму. Синергічну позитивну дію вони чинять на процеси глікогенолізу та секрецію α-амілази. Прикладом антагоністичної дії є їх вплив на інтенсивність серцевих скорочень: цАМФ стимулює, а цГМФ гальмує цей процес. Вважають, що реалізація всіх різноманітних дій Ц.н. забезпечується посередниками (месенджерами) третього порядку, роль яких виконують іони кальцію.
Биохимия / Под ред. Е.С. Северина. — М., 2003; Боєчко Ф.Ф., Боєчко Л.О. Основні біохімічні поняття, визначення і терміни. — К., 1993; Вороніна Л.М., Десенко В.Ф., Мадієвська Н.М. та ін. Біологічна хімія. — Х., 2000.