
РЕПЛІКАЦІЯ (лат. replicatio < replicare — відбивати) — процес самовідтворення молекул нуклеїнових кислот шляхом копіювання, передавання інформації в основному від ДНК до ДНК (процес подвоєння молекул ДНК) або в деяких вірусів від РНК до РНК. Р. відбувається за напівконсервативним механізмом, який вперше був запропонований Дж. Уотсоном та Ф. Кріком і доведений дослідами М. Меселсона і Ф. Сталя. Згідно з цим механізмом материнська молекула ДНК у процесі Р. розділяється на два ланцюги, кожен з яких служить матрицею для синтезу нового комплементарного ланцюга. Новоутворена молекула ДНК має один ланцюг вихідної ДНК (материнської) та один синтезований ланцюг (дочірній). Первинна структура дочірнього ланцюга визначається первинною структурою материнського ланцюга, тому що в основі її утворення лежить принцип комплементарності основ (А=Т і Г≡Ц). Це забезпечує видоспецифічне передавання генетичної інформації від покоління до покоління. Процес Р. складається з трьох основних стадій: утворення реплікативної вилки (ініціація), синтезу нових ланцюгів (елонгація), завершення синтезу двох дочірніх ланцюгів ДНК (термінація). Кожна стадія відбувається за участю відповідних ферментів і білкових факторів. Усього в процесі Р. беруть участь понад 20 різних ферментів і білків, кожен з яких виконує певну функцію. Весь комплекс називають ДНК-репліказною системою, або реплісомою. Для забезпечення напівконсервативного механізму Р. необхідним є розкручування подвійної спіралі ДНК та утворення одноланцюгової матриці. На початку Р. (стадія ініціації) на певних ділянках (точки початку Р., або точки ori — англ. origin — початок) відбувається локальна денатурація ДНК, ланцюги розходяться й утворюються реплікативні вилки (ділянки з V-подібною конфігурацією; рисунок).
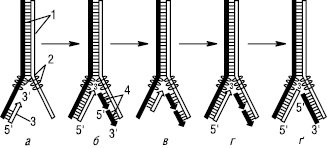
Рисунок. Механізм реплікації ДНК: а — розходження ланцюгів материнської ДНК і синтез РНК-затравки; б — утворення комплементарної гібридної ланки РНК-ДНК і синтез фрагментів Оказакі в напрямку 5 → 3´; в — гідроліз РНК-затравки; г — добудовування комплементарного ланцюга ДНК на місці РНК-затравки; ґ — зшивання фрагментів комплементарного ланцюга ДНК; де 1 — материнські ланцюги; 2 — SSB-білки; 3 — РНК; 4 — фрагменти Оказакі
Р. ДНК еукаріотів починається одночасно у багатьох місцях; вважають, що число точок початку реплікації перевищує тисячу. Із кожної точки одночасно у протилежних напрямках рухаються дві реплікативні вилки. Розкручування подвійної спіралі ДНК та відокремлення двох її ланцюгів відбувається при дії відповідних ферментів і білкових факторів. Так, ферменти ДНК-топоізомерази (І, ІІ і ІІІ) мають нуклеазну активність і регулюють суперспіралізацію ДНК. Напр., ДНК-топоізомераза І розриває фосфодіефірний зв’язок в одному з ланцюгів подвійної спіралі й ковалентно приєднується до 5′-кінця у точці розриву. Після закінчення формування реплікативної вилки фермент ліквідує розрив у ланцюзі й відділяється від ДНК. Ферменти хелікази (лат. helix — спіраль) розплітають короткі ділянки ДНК, які знаходяться безпосередньо перед реплікативною вилкою. Для розділення кожної пари основ витрачається енергія гідролізу двох молекул АТФ. Унаслідок цього відбувається розкручування ділянки суперспіралізованої молекули ДНК. У підтриманні цієї ділянки ДНК у розкрученому стані беруть участь SSB-білки (англ. single strand binding proteins), які зв’язують одноланцюгові нитки ДНК, протидіючи їх повторному об’єднанню. Внаслідок багатоцентрової Р. хромосоми повне подвоєння генетичного матеріалу клітин вищих організмів (зокрема ссавців) відбувається протягом 9 год. Після розкручування подвійної спіралі ДНК утворюються дві одноланцюгові матриці, один ланцюг при цьому має напрямок 3′→5′, а другий — 5′→3′ (оскільки у дволанцюговій молекулі ДНК ланцюги антипаралельні). Субстратами та джерелами енергії для синтезу нового ланцюга служать 4 дезоксирибонуклеозидтрифосфати — дАТФ, дГТФ, дЦТФ і дТТФ, від яких відщеплюється по пірофосфатному залишку, що й забезпечує енергією утворення кожного нового фосфодіефірного зв’язку. Синтез нових ланцюгів (елонгація) відбувається тільки в напрямку 5′→3′, антипаралельно до матричного ланцюга. Напрямок одного ланцюга збігається з напрямком руху реплікативної вилки (лідируючий ланцюг), а інший проходитиме в протилежному напрямку (відстаючий ланцюг). Тому провідний ланцюг синтезується безперервно в напрямку руху реплікативної вилки, а відстаючий — переривчасто з утворенням фрагментів, які потім зшиваються. Ці фрагменти одержали назву фрагментів Оказакі (за прізвищем ученого, який їх відкрив у 1968 р.). Фрагменти Оказакі синтезуються у напрямку, протилежному руху реплікативної вилки. Їх довжина у прокаріотів становить 1000–2000 нуклеотидів, а в еукаріотів значно менша (≈100 нуклеотидів). Р. ДНК здійснюється ДНК-залежними ДНК-полімеразами. Клітини у прокаріотів та еукаріотів містять декілька ДНК-полімераз. У прокаріотів у процесі Р. беруть участь ДНК-полімерази І, ІІ і ІІІ. У синтезі еукаріотичних ДНК беруть участь 5 ДНК-полімераз — α, β, γ, δ, ε. Головний фермент, що каталізує Р. обох ланцюгів у прокаріотів, — ДНК-полімераза ІІІ, а в еукаріотів — ДНК-полімераза α. Інші ДНК-полімерази відіграють допоміжну роль у Р. Показано, що ДНК-полімерази не спроможні почати синтез нового ланцюга, а для ініціації синтезу ДНК необхідні короткі (10–200 нуклеотидів) послідовності РНК, які виконують роль приманок (праймерів) і синтезуються за допомогою специфічної РНК-полімерази (праймази або примази). Синтез фрагментів Оказакі також починається з праймера, який після завершення синтезу фрагментів видаляється, а розриви між фрагментами заповнюються дезоксирибонуклеотидами під дією ДНК-полімерази І. Врешті фрагменти Оказакі з’єднуються один з одним за допомогою ферменту ДНК-лігази. Два нові ланцюги, з’єднані зі своїми комплементарними ланцюгами, утворюють дві дочірні подвійні спіралі, кожна з яких містить один материнський і один новосинтезований ланцюг. Важливе значення має здатність ДНК-полімераз проявляти, крім полімеразної, ще й екзонуклеазну активність. Якщо в ході Р. включиться некомплементарний нуклеотид, фермент робить крок назад, відщеплює неправильний нуклеотид і після цього знову продовжує полімеразні реакції. Завдяки такому коригуванню частота помилок при Р. ДНК не перевищує 1 на 109–1010 нуклеотидів.
Після завершення Р. відбувається метилування нуклеотидних залишків синтезованих ланцюгів ДНК. Метильні групи приєднуються до всіх залишків аденіну в послідовності — ГАТЦ — з утворенням N6-метиладеніну, а також можливе утворення N5-метилцитозину. Кількість метильованих основ становить 1–8%. Наявність метильних груп у ланцюгах ДНК необхідна для формування структури хромосом, а також для регуляції транскрипції генів. На кожному кінці хромосоми присутня специфічна нуклеотидна послідовність — багаточисленні повтори (сотні або тисячі разів) олігонуклеотидів — ГГГТТА, — які називають теломірною послідовністю чи просто теломірною ДНК. Наявність теломірів необхідна для завершення Р. кінцевих інформативних послідовностей хромосом, тобто для збереження генетичної інформації. На процес Р. можуть діяти різноманітні речовини, у т.ч. антибіотики. Їх широко використовують у біохімічних дослідженнях. Так, мітоміцин С утворює ковалентні зшивки між двома комплементарними ланцюгами ДНК і перешкоджає їх розходженню, гальмуючи процес Р. Цей антибіотик виявляє антиканцерогенну дію, блокуючи поділ пухлинних клітин.
Биохимия / Под ред. Е.С. Северина. — М., 2003; Гонський Я.І., Максимчук Т.П. Біохімія людини. — Тернопіль, 2001; Ленинджер Л. Основы биохимии: В 3 т. — М., 1985. — Т. 3.